| |
 |
THE OYSTERCATCHER SHELL GAME
Hybrid or frazari? Does it matter?
|
 An interesting white-bellied oystercatcher first found by Bonnie Burgess on 27 Feb 2021 at Yankee Pt., has sparked much local interest in oystercatcher identification (photo right © B. Burgess). It remained with Black Oystercatchers Haematopus bachmani ["BLOY" often hereafter] at Yankee Pt. through the morning of 9 March, was refound at Otter Pt., Pacific Grove by Diane Bell on 10 March, and then frequented the rocky shore at Asilomar SB, Pacific Grove, from at least 12-14 March (photo above © John Drum). It is an adult and the split iris indicates it is a female (fide Rick Hanks, director of the California Central Coast BLOY Project for the Monterey Bay region). When sitting or foraging, it strongly resembled an American Oystercatcher H. palliatus [sometimes "AMOY" hereafter] with its red bill, black head, brown back, pink legs, and white belly. In flight, however, it was apparent that it had white wingstripe restricted to the greater secondary coverts, and dusky smudging on the flanks, axillaries, and uppertail coverts. The issue raised was is this a vagrant of the west Mexican race H. p. frazari [just "frazari" hereafter] or did these latter characters suggest it was likely a hybrid between frazari and a BLOY [i.e., American Oystercatcher X Black Oystercatcher in eBird lingo]? An interesting white-bellied oystercatcher first found by Bonnie Burgess on 27 Feb 2021 at Yankee Pt., has sparked much local interest in oystercatcher identification (photo right © B. Burgess). It remained with Black Oystercatchers Haematopus bachmani ["BLOY" often hereafter] at Yankee Pt. through the morning of 9 March, was refound at Otter Pt., Pacific Grove by Diane Bell on 10 March, and then frequented the rocky shore at Asilomar SB, Pacific Grove, from at least 12-14 March (photo above © John Drum). It is an adult and the split iris indicates it is a female (fide Rick Hanks, director of the California Central Coast BLOY Project for the Monterey Bay region). When sitting or foraging, it strongly resembled an American Oystercatcher H. palliatus [sometimes "AMOY" hereafter] with its red bill, black head, brown back, pink legs, and white belly. In flight, however, it was apparent that it had white wingstripe restricted to the greater secondary coverts, and dusky smudging on the flanks, axillaries, and uppertail coverts. The issue raised was is this a vagrant of the west Mexican race H. p. frazari [just "frazari" hereafter] or did these latter characters suggest it was likely a hybrid between frazari and a BLOY [i.e., American Oystercatcher X Black Oystercatcher in eBird lingo]? |
 I'd like to give a different perspective on this topic by reviewing the literature, especially Jehl (1985), and questioning some of our assumptions. The photo (left) is from Jehl (p. 495) showing a mixed pair of oystercatchers from Scammons Lagoon, Baja California. The purpose of his paper "is to clarify species limits by documenting the nature of variation in size and plumage among populations of oystercatchers on the west coast of North and Middle America, with special reference to the zone of overlap." I'd like to give a different perspective on this topic by reviewing the literature, especially Jehl (1985), and questioning some of our assumptions. The photo (left) is from Jehl (p. 495) showing a mixed pair of oystercatchers from Scammons Lagoon, Baja California. The purpose of his paper "is to clarify species limits by documenting the nature of variation in size and plumage among populations of oystercatchers on the west coast of North and Middle America, with special reference to the zone of overlap."
The population of American Oystercatcher in Baja California and west Mexico is "Frazar's Oystercatcher" H. p. frazari. Jehl created a "character index" in which a "pure" Black Oystercatcher would score 0 and an American Oystercatcher in eastern North America [H.p. palliatus] would score 42. Ten plumage characters would be scored from 0-5 or 0-7 "to classify the range of variation." To represent BLOY, Jehl chose his scores of birds from Oregon northwards; to represent AMOY he used "populations of frazari in the northern Gulf of California and the coasts of Sonora and Sinaloa" (p. 490). Using just that set of AMOY, he proposed to "consider character scores of 30-38 (mean of 33.6) as typical of frazari. For purposes of this paper, any individual with a score of 10-29 is considered to be a hybrid;" (p. 491).
The first assumption I wish to disavow is that Jehl's character index, available on-line, was meant to define what was actually a hybrid or not. He had no such intent — indeed, in 1985, he had no genetic information at all. Nor is the Jehl score meant to categorize the birds of western Baja California. He intentionally used scores from specimens far from the area of species' overlap to use as a tool to analyze what he found in the overlap zone. The Jehl score is not an evaluation of Baja oystercatchers, and the borders drawn within the index (e.g., 10-29 as hybrid) is not meant to identify hybrids. Still, it is a very nice tool to evaluate "color variation as expressed in the degree to which dark coloration invades the white areas" of AMOY (p. 499). |
GLOSSARY for this page
Hybrid swarms occur when the hybrid is viable and at least as vigorous as its parent types; and there are no barriers to crossbreeding between the hybrid and parent types. Swarms cannot occur if one of these conditions is not met (Ward et al. 2012).
An F1 Hybrid is the first filial generation of offspring of distinctly different parental types. F1 hybrids are used in genetics. The term is sometimes written with a subscript for the "1". |
Speciation via hybridization and introgression: Species may evolve in sympatry and in isolation; climate change and the movement of land by tectonic plates contribute to isolation and the formation of new terrestrial species, but introgressive hybridization can be a factor. "Exaggerated plumage traits (crest and tail length) have entered populations of two species of paradise flycatchers (genus Tersiphone) as a result of hybridization with a third species, in both east and west Africa. . . . The relevance to speciation lies in the fact that regions of introgression are peripheral areas, which could become isolated from the main range of the species through a change in climate and habitat: they are potential sites of speciation;" Grant & Grant (1997; derived in part from their work on Galapagos finches).
"After a long period of geographic isolation, the cause of which may be either natural or man-made, the re-occurrence of the parent species in the same environment can lead to interbreeding, hybridization, and potentially, a hybrid swarm. Hybrid swarms can lead to introgressive hybridization, or introgression, in which there is gene flow from the hybrid gene pool to the parental gene pool, or vice versa, occurring due to backcrossing between the populations;" Harrison et al. (2014). |
|
Direct hybridization (i.e., F1 hybrids) can lead to multiple back-crossing introgression, and the flow of genes expressing such introgression, through a population of organisms. In birds it is one method by which new species develop (Grant & Grant 1997). It is neither a good thing or a bad thing — it is a normal part of evolution as an organism adapts to environmental changes. Some introgression that occurred eons ago may remain in the organism's genome. In humans, up to 4% of the genome contains Neanderthal ancestry as a direct result of admixture with these archaic species (Green et al. 2010). As Neanderthals went extinct about 50,000 years ago, that is surely very ancient hybridization [no jokes here about your ability to pick out those people currently with the most Neanderthal genes].
The parasitic blood fluke Schistosoma haematobium, the one most commonly the cause of the disease schistosomiasis in humans, may hybridize with livestock schistosomes, including S. bovis. A recent study in Niger and Zanzibar found no evidence for contemporary hybridization between S. bovis and S. haematobium in recent samples. However, all S. haematobium genomes from Niger revealed genetic evidence of an ancient introgression event that occurred at least 108–613 generations ago; Pratt et al. (2019). These worms live 3-5 years on average, and lay eggs throughout their lifespan. I'm not sure how long in time 108-613 generations would take.
 We know that oystercatchers are long-lived, have slow reproductive rates, and have low annual productivity success, often below 20% (i.e., reports of the California Central Coast BLOY Project for the Monterey Bay region [fide H. Hanks]). Jehl (1985) reported that "some examples of H. unicolor (New Zealand; photo right), H.p. durnfordi (Argentina), and H.p. galapagensis (Galapagos Is.) may be distinguishable from frazari only on the basis of locality;" (p. 499). From this fact he deduced that each population had gone through ancient periods of hybrid swarms, or in the case of galapagensis, a Middle American ancestor had been "derived from a hybrid population" (p. 499). We know that oystercatchers are long-lived, have slow reproductive rates, and have low annual productivity success, often below 20% (i.e., reports of the California Central Coast BLOY Project for the Monterey Bay region [fide H. Hanks]). Jehl (1985) reported that "some examples of H. unicolor (New Zealand; photo right), H.p. durnfordi (Argentina), and H.p. galapagensis (Galapagos Is.) may be distinguishable from frazari only on the basis of locality;" (p. 499). From this fact he deduced that each population had gone through ancient periods of hybrid swarms, or in the case of galapagensis, a Middle American ancestor had been "derived from a hybrid population" (p. 499).
My point is that ancient hybridization and introgression, occurring millions of generations ago, can be expressed currently from the archaic genes in the genome. For the Galapagos AMOY, that must have occurred more than 3-5 million years ago, as the islands are geologically "new" and didn't appear until about then. The Baja Peninsula split from the mainland about 12-15 mya, providing an overlap with BLOY for millions of years. We cannot assume that the current plumage of any oystercatcher represents recent genetic hybridization — it could well be an expression of ancient hybrids. Surely there is something like a "statute of limitations" to be applied to archaic expressions of long-distance gene flow? |
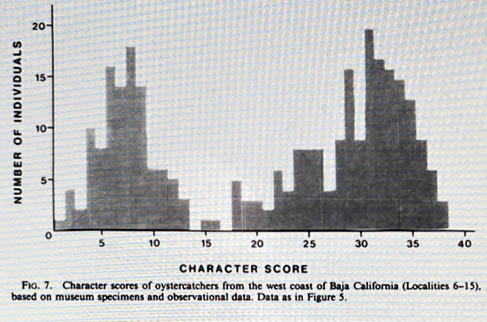
The results of Jehl's study are summarized in the Abstract to his paper (below left), and shown nicely in figure 7 (below right, from p. 497)
| The ranges of the Black Oystercatcher (Haematopus bachmani) and the American Oystercatcher (Haematopus palliatus frazari) overlap for approximately 480 km along the Pacific coast of Baja California. Phenotypes are variable, especially in coloration, and hybridization has long been known. Yet, at most localities parental morphs predominate, hybrids are uncommon, and assortative mating occurs when possible. The historical record is sufficiently detailed to allow demonstration of changes in the hybrid zone. At the turn of the century — or whenever they were first studied — most populations were composed largely of parental morphs. After intensive collecting, resulting in the virtual extermination of some populations, recolonization occurred from both north and south. The reconstituted populations differed from the original ones, and by the 1920s and 1930s the frequency of hybridization was high. Recently, however, many populations have returned to their original composition. The prevalence of parental forms, a demonstration of assortative mating, and the resumption of stability in the zone of hybridization after a period of dynamic change, all show that there is selection against hybridization in this zone of secondary contact and that the two forms are specifically distinct. The nature of the selective forces remains to be determined. |
 |
|
Figure 7 shows the scores of about 270 oystercatchers within the range of BLOY-AMOY overlap, which is approximately the northern half (300 miles) of the Baja Peninsula. Jehl (pp. 497-498) says the figure provides evidence of mate preference in the zone of overlap: "If matings occurred at random, the result would be a hybrid swarm and the distribution on character scores would approximate a normal distribution [meaning, a standard bell curve graph]. The opposite is the case (fig. 7). The distribution is strongly bimodal, clustering near "pure" scores for each of the parental forms; intermediate scores are poorly represented. I interpret this to mean that assortative mating is prevalent" [emphasis added].
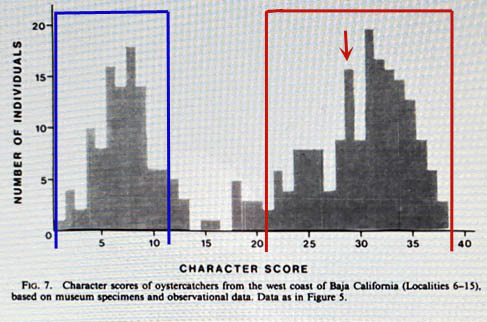 You will recall that Jehl's character index, established from just northern Gulf of California AMOYs, had a mean of 33.6. Later (p. 499), he states that if scores from southwestern Baja to southern Mexico are included, the "mean scores vary from 28.3 to 38.6 [overall range 20-42], suggestive of hybridization and introgression with bachmani to the north and palliatus to the south." Those areas are all outside the overlap zone in northern Baja, where there was active hybridization in the 1920-1930s as populations from the north and south took over empty territories after the extirpation of oystercatchers from that zone. Thus, such a revised index would include expressions of ancient hybridization but exclude those from 100 years ago. I've moved the "mean" on a revised figure 7 [right] with a new AMOY mean set at 28 (red arrow) and a bell curve distribution for each bimodal distribution (blue box for BLOY, red box for AMOY. I suggest that these boxes better represent an index for hybridization than the Jehl chart comparing Gulf of California birds to overlap zone. You will recall that Jehl's character index, established from just northern Gulf of California AMOYs, had a mean of 33.6. Later (p. 499), he states that if scores from southwestern Baja to southern Mexico are included, the "mean scores vary from 28.3 to 38.6 [overall range 20-42], suggestive of hybridization and introgression with bachmani to the north and palliatus to the south." Those areas are all outside the overlap zone in northern Baja, where there was active hybridization in the 1920-1930s as populations from the north and south took over empty territories after the extirpation of oystercatchers from that zone. Thus, such a revised index would include expressions of ancient hybridization but exclude those from 100 years ago. I've moved the "mean" on a revised figure 7 [right] with a new AMOY mean set at 28 (red arrow) and a bell curve distribution for each bimodal distribution (blue box for BLOY, red box for AMOY. I suggest that these boxes better represent an index for hybridization than the Jehl chart comparing Gulf of California birds to overlap zone.
|
There is yet another way to look at the concept of restricting our review of current birds to current hybridization patterns. Jehl (1985) found that the current situation, and presumably the "original" situation in the 19th century before collecting essentially extirpated oystercatchers in northern Baja, was a "high frequency of assortative mating ... as well as a low frequency of hybrids" (p. 499). The CBRC's book on rare birds in California (CBRC 2007), including Appendix H and on-line reports through 2008, indicates that about 100 pied oystercatchers have been accepted, rejected, or were reported but unsubmitted because of the hybrid issue. Of those 39% were accepted (returning birds are not double-counted) and 61% rejected. Those statistics suggest that about 60% of the vagrants from Baja are hybrids. But that is statistically improbable given Jehl's finding of a "low frequency of hybrids" within the overlap zone. In fact, looking at the red box around Figure 7, and knowing that that mean (red arrow) is a score of 28, if one "accepted" as "pure" only those scoring 30 or higher, you would effectively reject about 60% of the birds in that distribution.
In short, by using a score of 30 as a cut-off, the scoring becomes a means of choosing as "pure" only the whitest end of the AMOY cline. There is something unsettling about that approach, as Jehl's interpretation of history was that all of the subspecies frazari "originated in hybrid swarms in zones of secondary contact." I think it is wrong to assume there are any "pure" frazari — meaning those without any ancient or archaic hybridization in their genome. Since we have no genetic evidence on this topic, it seems rather pointless to me to "accept" the whitest end of a distribution of character types. We have zero evidence of any proven hybrids within the red-boxed set of distributions. The pattern of color distribution found by Jehl could all be an expression of genetic patterns years ago as ice-ages and sea-level rise created scarcity of breeding habitat. As Jehl further noted (p. 495): "because populations are small and stable, an unmated bird may have to accept (not select) any bird of the opposite sex that becomes available if it is to enter the breeding population. Thus, the existence of mixed pairs is not necessarily evidence for random mating, or, more precisely, evidence against a preference for assortative mating."
 "Frazar's Oystercatcher" was described by Brewster (1888) from specimens collected in the Gulf of California. He described the "species" as differing from nominate palliatus in, to quote Jehl's rendition (p. 488), "being slightly browner dorsally, having the white areas invaded to a variable degree by dark smudging, especially on the rump, undertail coverts, wing lining, and margin of chest band; in addition, the amount of white on the primaries is reduced." It was actually Murphy (1925) who noted the wingstripe, with white spots on the primaries "wholly lacking" in most specimens. Murphy (1925) further stated that "in its maximum expression, when the whole breast, sides, flanks, and undertail coverts are heavily blotched, the character is peculiar to frazari." Don't these descriptions of 19th century specimens describe the current Monterey bird well? See Bill Hill's photo (left). "Frazar's Oystercatcher" was described by Brewster (1888) from specimens collected in the Gulf of California. He described the "species" as differing from nominate palliatus in, to quote Jehl's rendition (p. 488), "being slightly browner dorsally, having the white areas invaded to a variable degree by dark smudging, especially on the rump, undertail coverts, wing lining, and margin of chest band; in addition, the amount of white on the primaries is reduced." It was actually Murphy (1925) who noted the wingstripe, with white spots on the primaries "wholly lacking" in most specimens. Murphy (1925) further stated that "in its maximum expression, when the whole breast, sides, flanks, and undertail coverts are heavily blotched, the character is peculiar to frazari." Don't these descriptions of 19th century specimens describe the current Monterey bird well? See Bill Hill's photo (left).
In addition to this discussion, we know that Jehl himself approved, as a second California record, a specimen with a score more consistent with the red box concept outline above, than his "character index." As it happens, I wrote the CBRC report (Roberson 1993) that commented on the acceptance of the State's first two records: |
One collected at San Diego, SD, 16 May 1862 (MVZ 4488; 59-1985) and another collected on Santa Barbara I., SBA, 2 June 1863 (MVZ 4489), both collected by W. Cooper.
Both birds provoked questions about possible hybridization with the Black Oystercatcher (H. bachmani). Jehl (1985) devised a scoring system ranking ten characters (e.g., color of uppertail coverts, extent of white wing stripe) on a scale from 0 to 4 or 0 to 6. Under this system, scores below 10 specify H. bachmani, those above 30 H. p. frazari, and those between suggest introgression. Joseph Morlan and John Trochet studied the specimens and scored them independently; Joseph R. Jehl, Jr., also provided his original scores of these two specimens. The San Diego bird was scored 35 by Jehl, 35 by Morlan, and 33 by Trochet; it is clearly an American Oystercatcher. The Santa Barbara Island bird was scored 27 by Jehl, about 29 by Morlan, and about 26 by Trochet, scores borderline and problematic. Jehl considers the bird to be frazari. Jehl (1985) hypothesized that widespread collecting around the turn of the century on the islands off Baja California led to hybridization; Cooper's birds were taken prior that collecting. |
|
|
 It is apparent in my comments in 1993 that, at that time, I was a fan of Jehl's character index, and possibly thought it actually indicated what birds were likely to be hybrids. Yet he scored the specimen — which is much easier than scoring photos — at 27. His acceptance suggests, to me, that his "character index" border at 30 was meant for analyzing his Baja data for his 1985 paper, and not an effort to try to identify "pure" frazari, whatever that may mean. It is apparent in my comments in 1993 that, at that time, I was a fan of Jehl's character index, and possibly thought it actually indicated what birds were likely to be hybrids. Yet he scored the specimen — which is much easier than scoring photos — at 27. His acceptance suggests, to me, that his "character index" border at 30 was meant for analyzing his Baja data for his 1985 paper, and not an effort to try to identify "pure" frazari, whatever that may mean.
The current Monterey County bird (right) has been independently scored six times: 3 by observers (including a scoring that I did with my wife Rita Carratello), and 3 outside experts who used a wide selection of photos. The scores were (low to high): 22, 24, 23-25, 26.5, 29, 29. Let's throw out the low score and one high score (i.e., we'll throw out my score), and let's restate the score that has a range (23-25) as 24, and the one with a partial score (26.5) to adjust downwards to this set of four: 24, 24, 26, 29. The average of those scores is 25.7. It is on the darker side of the ledger but is within about 2.5 points of Jehl's lowest mean (28.3) for frazari's full range (excluding northern Baja). The most prominent differences among scorers related to width of wing stripe, thighs, and undertail coverts — parts of the anatomy that can be hard to review in photographs — and further illustrate why the Jehl scale was originally intended to apply primarily to specimen review.
|
About midway down the Baja Peninsula "the predominantly rocky shores and islands of western North America give way to the light colored sandy beaches of southern Baja California. It is in this region that the ranges of the two oystercatchers overlap;" (Jehl, p. 491). There are many barriers to producing hybrid young in today's conditions. Oystercatchers lay eggs in nests on the ground. To avoid mammalian predators, nearly all nests are on islands or islets off the mainland.
 Jehl explains some problems faced: "First, the number of nesting sites on some islands is limited, oystercatchers are long-lived, and pairs remain intact for many seasons, perhaps for life. Thus, it is probably difficult for young birds to gain a territory and to enter the breeding population near their natal site. Jehl explains some problems faced: "First, the number of nesting sites on some islands is limited, oystercatchers are long-lived, and pairs remain intact for many seasons, perhaps for life. Thus, it is probably difficult for young birds to gain a territory and to enter the breeding population near their natal site.
Second, because the reproductive life of an oystercatcher is very long, it follows that annual production of breeders or survivorship of non-breeders is low. And, because pair bonds are persistent, generation times are long, the opportunities for backcrossing and introgression are reduced, and selection to establish or reinforce potential isolating mechanism acts slowly. . .
[In addition], there is a general correlation between substrate color and adult phenotypes, although it is obscured on [some islands]. Oystercatcher chicks are polymorphic, being dark-colored in bachmani and pale with dark marking in frazari (Jehl 1968). In broods of mixed parentage some chicks will not be cryptically colored (see Bancroft 1927). The probability that those will be quickly discovered by Western Gulls (Larus occidentalis) seems high;" (pp. 497-498).
Copied here is the photo in Bancroft (1927), showing two fledglings from one nest at Scammons Lagoon in 1926. He did not state the plumages of the parents, but these fledglings may well by F1 hybrids. After the photo, Bancroft collected these young.
|
So, in summary, my conclusion are:
- Isn't this surely an obvious hybrid from the "hybrid swarm" of oystercatchers in Baja California?
Answer: No, there is no "hybrid swarm" at any recent time in Baja California. The most recent "swarm" was about 100 years ago. To quote Jehl's response to that question: "The opposite is the case" (Jehl 1985, p. 498).
- Can't one just use the "Jehl index," get a score, and label a specific bird by its score?
Answer: You may, but the Jehl index was not devised for that purpose; was not devised from specimens in Baja California so does not address their plumage distribution; is quite subjective to use; and Jehl himself identified a specimen as an acceptable frazari to establish California's second record, despite a score in the "hybrid" zone of his index.
- Can't one classify a "pure" frazari from a high score on the Jehl index?
Answer: Probably not. Jehl opined (p. 499) that "frazari [and several subspecies elsewhere in the world] are not classical subspecies but originated as hybrid swarms in zones of secondary contact." Even the race galapagensis, isolated in the Galapagos Islands, may have "shared a common ancestor [with frazari] and that the Galapagos residents were derived from a hybrid population in Middle America." Unless one finds a way to ignore ancient hybridization within a genome, there are no "pure" birds in any of these populations, including frazari.
- Is the Monterey bird a hybrid?
Answer: Maybe, we don't know. There is no genetic evidence one way or the other. It is probably not a recent hybrid, and is unlikely to be an F1 hybrid or the result of any recent mating. It shows many characters that have long been embedded in the frazari genome, which establishes genetic introgression at some point in the past in this bird's family history, but that could have been hundreds or thousands of years ago. If your preferred definition of a "hybrid" includes ancient hybridization stretching back into millennia, you will prefer to call it a "hybrid." In fact, as discussed above, you should probably consider the entire oystercatcher population within the range of frazari as "hybrids." By that criteria, no pied oystercatcher from Baja should ever be accepted in California.
- Is the Monterey bird a frazari American Oystercatcher?
Answer: It is surely an oystercatcher hatched within the known range of frazari, and very likely from Baja California. Whatever it is, it is a rare vagrant to central California. Jehl found that in the zone of overlap in northern Baja, and after decades to recover from the over-collecting there, there was a "high frequency of assortative mating ... as well as a low frequency of hybrids." I think the Monterey bird's plumage score likely expresses the standard frazari genome with its long history of archaic hybridization. Because hybrids are scarce, and there are many barriers to produce a hybrid, the rule of parsimony suggests to me that this is most likely a normal darkish-end frazari. The likelihood of it being a recent hybrid is comparatively small.
So what do you do when a named taxon, defined by a suite of characters that suggest a long history of hybridization in the distant past, goes through another period of active hybridization a hundred years ago, but has re-established assortative mating and isolating mechanisms that select against hybrids? What if the characters that might suggest recent hybridization are the same characters for which the taxon was defined centuries ago? I suggest that the most reasonable approach is to accept "it is what it is." One would try to weed out F1 hybrids or clear intermediates (e.g., an all black oystercatcher with white vent and partial white wingstripe; there are photos of birds like this on-line from Baja). Otherwise, if the oystercatcher is clearly on the AMOY side of the ledger, with dark smudging invading the white areas "to a variable degree, especially on the rump, undertail coverts, wing lining, margin of chest band . . and having the amount of white on the primaries reduced or absent," as described by Brewster (1888) and Murphy (1925), I'd identify it as frazari and be content. The amazing fact is that it reached California from a distant and definable population. I don't see the point in trying to select some "whitest" end of a cline described long ago.
I was going to discuss the 11 extant species oystercatchers (6 pied, 4 black, and one with pied and black morphs), but this is too long already. You can find a review of the Oystercatcher family [Haematopodidae] on this web site.
Finally, for fun, here are two famous California records of American Oystercatcher. First, an adult frazari on Anacapa Island, Ventura Co., present from 24 May 1964–19 Oct 1980 (below left; photo in spring 1975 © Dave Rudholm), that remained for 16 years (!) and represents the long-staying of all State AMOYs. Second, a photo of two of 3 juv American Oystercatchers at Salton City, Imperial Co., from 14-19 Aug 1977, and then at Salt Creek, Riverside Co., from 20-30 Aug 1977 (below right, photo © Jeri M. Langham), that are the only interior record for the State. Patten et al. (2003) reviewed photos of the Salton Sea birds, and suggested that these birds may have been nominate H. p. palliatus, perhaps from the Gulf of Mexico. If all California records of frazari were rejected on grounds of ancient hybridization, these would represent the only State record. |
|
| |
Acknowledgments: I thank Curtis Marantz for providing a copy of Jehl (1985) for this study. I'm grateful to those who provided expert comments, discussion, or scoring of oystercatchers for this project — thanks to Bonnie Burgess, Rita Carratello, Curtis Marantz, Joe Morlan, Andrew Spencer, and Brian Sullivan. Rita Carratello and Michael Rieser graciously read an earlier version of this page, and suggested many corrections. I thank Jeri Langham and Dave Rudholm for the old CA photos, and thank all the photographers of the Monterey individual. Some particularly useful photos were provided by Bonnie Burgess, Terence Degan, John Drum, Bill Hill, Carole Rose and Brian Sullivan; quite a number of these are posted in eBird. I also reviewed photos from eBird of oystercatchers in Baja California and Baja California Sur, and appreciate the efforts of those photographers. The opinions expressed here are mine, and not necessarily shared by those acknowledged here. There is a wide variety of opinions on these topics.
Literature cited or consulted:
Bancroft, G. 1927. Breeding birds of Scammons Lagoon. Condor 29: 29-53.
Brewster, W. 1888. Descriptions of supposed new birds from Lower California, Sonora, and Chihuahua, Mexico, and the Bahamas. Auk 5: 82-95.
California Bird Records Committee (R.A. Hamilton, M.A. Patten, and R.A. Erickson, eds). 2007. Rare Birds of California. West. Field Ornithologists, Camarillo, CA.
del Hoyo, J., and N.J. Collar. 2014. HBW and BirdLife Intern. Illustrated Checklist of the Birds of the World, Vol. 1: Non-passerines. Lynx, Barcelona.
Dickinson, E.C., and J.V. Remsen, Jr., eds. 2013. The Howard & Moore Complete Checklist of the Birds of the World. 4th ed, Vol. 1. Aves Press, Eastbourne, U.K.
Grant, P.R., and B.R. Grant. 1997. Genetics and the origin of bird species. Proc. Nat. Acad. Sci. 94: 7768–7775.
Green RE, J. Krause, A.W. Briggs, T. Maricic, U. Stenzel, M. Kircher, N. Patterson, H. Li, W. Zhai, MH-Y Fritz, et al. 2010. A draft sequence of the Neanderthal genome. Science 328:710–722.
Harrison, R.G., and E.L. Larson. 2014. Hybridization, Introgression, and the Nature of Species Boundaries. J. Heredity. 105: 795–809.
Jehl, J., Jr. 1968. Relationships in the Charadrii (shorebirds: a taxonomic study based on color patterns of the downy young. San Diego Soc. Nat. Hist. Mem. 3.
Jehl, J.R., Jr. 1985. Hybridization and evolution of oystercatchers on the Pacific Coast of Baja California. Ornith. Monogr. No. 36.
Murphy, R.C. 1925. Notes on certain species and races of oyster-catchers. Am. Mus. Novit. No. 194.
Roberson, D. 1993. Fourteenth report of the Calif. Bird Records Committee. Birds 24: 113-166.
Sibley, C.G., and B.L. Monroe, Jr. 1990. Distribution and Taxonomy of Birds of the World. Yale Univ. Press, New Haven, CT.
Patten, M.A., G. McCaskie, and P. Unitt. 2003. Birds of the Salton Sea: Status, Biogregraphy, and Ecology. Univ. of Calif. Press, Berkeley.
Platt, R.N. II, M. McDew-White, W. Le Clec’h, F.D. Chevalier, F. Allan, A.M Emery, A. Garba, A.A. Hamidou, S.M. Ame, and J.P. Webster. 2019. Ancient Hybridization and Adaptive Introgression of an Invadolysin Gene in Schistosome Parasites. Molec. Bio. & Evol. 36: 2127–2142.
Ward, J.L., M.J. Blum, D.J. Walters, B.A. Porter, N. Burkhead, and B. Freeman. 2012. Discordant introgression in a rapidly expanding hybrid swarm. Evolutionary Applications 5: 380–392.
Winkler, D.W., S.M. Billerman, & I.J. Lovette. 2015. Bird Families of the World. Lynx Edicions, Barcelona.
Working Group, A. O., E. Nol, and R C. Humphrey. 2020. American Oystercatcher (Haematopus palliatus), version 1.0. In Birds of the World (A. F. Poole, Editor). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.ameoys.01
|
|
|

